
Type II Apoptosis
Macroautophagy or autophagy translates as 'self eating' which is an apoptotic process in response to nutrient starvation, protein aggregation, Endoplasmic Reticulum (ER) stress, calcium overload, hypoxia, removal of damaged organelles and proteasome impairment. Autophagy can be triggered as a cell surivial mechanism and hence may prevent cell death and thus it is integral to human health in terms of organism development, cell homestasis and is involved in a wide range of diseases including cancer, neurodegeneration, aging and the innate immune resposne to pathogens.
Autophagy was first fully described in 1963 by Christian de Duve who observed cytoplasmic material such as mitochondria and endoplasmic reticulum being engulfed by double-membrane vesicles or autophagosomes and digested by lysosomal enzymes after fusion with lysosomes.
Autophagy can be characterized morphologically in a similar manner to that of other forms of cell death by cell shrinkage, formation of autophagsomes and degradation of cell orgnalles. There are numerous signalling routes that regulate autophagy, these include mTOR and JNK signalling. Autophagy can be induced by nutrient starvation, formation of cytosolic protein aggregates, ER stress, calcium overload and damged organelles.
There numerous methods of inducing autophagy including serum and total nutrient starvation and rapamycin which both inhibit mTOR signalling. Chloroquine can also induce autophagy as well as ER-ATPase inhibitor, Thapsigargin which causes degradation of the ER.
Morphological Changes
- Cell shrinkage
- Double membrane vesicles or autophagosomes
- Degradation of organelles
- mTOR
- PI3 kinases
- UPR stress sensors
- Kinases - JNK
- Bcl-2
- IP3-receptor
Stimulus
- Nutrient starvation
- Protein aggregation
- Ischemia
- Hypoxia
- Calcium overload
- ER Stress
- Damaged organelles
- Proteasome impairment
Model of Autophagy
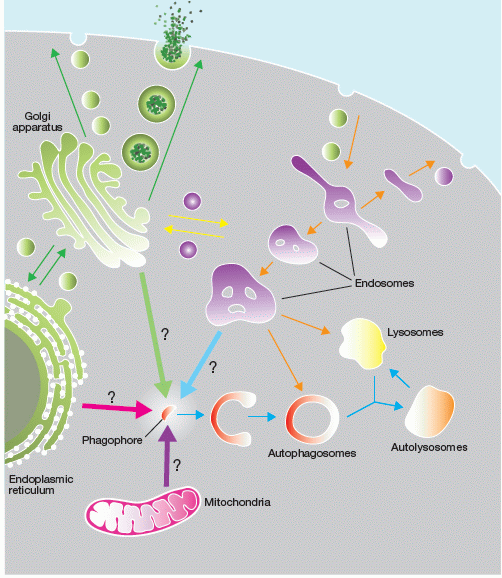
Figure by SA Tooze, The orgin of the autophagosomal membrane, Nature Cell Biology, 12, (9), p831-835, 2010.
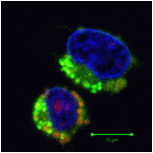
Autophagic LC3B expression
Role of Lysosomes in Autophagic Responses to Drugs
Role of Mitochondria in Autophagic Responses to Drugs
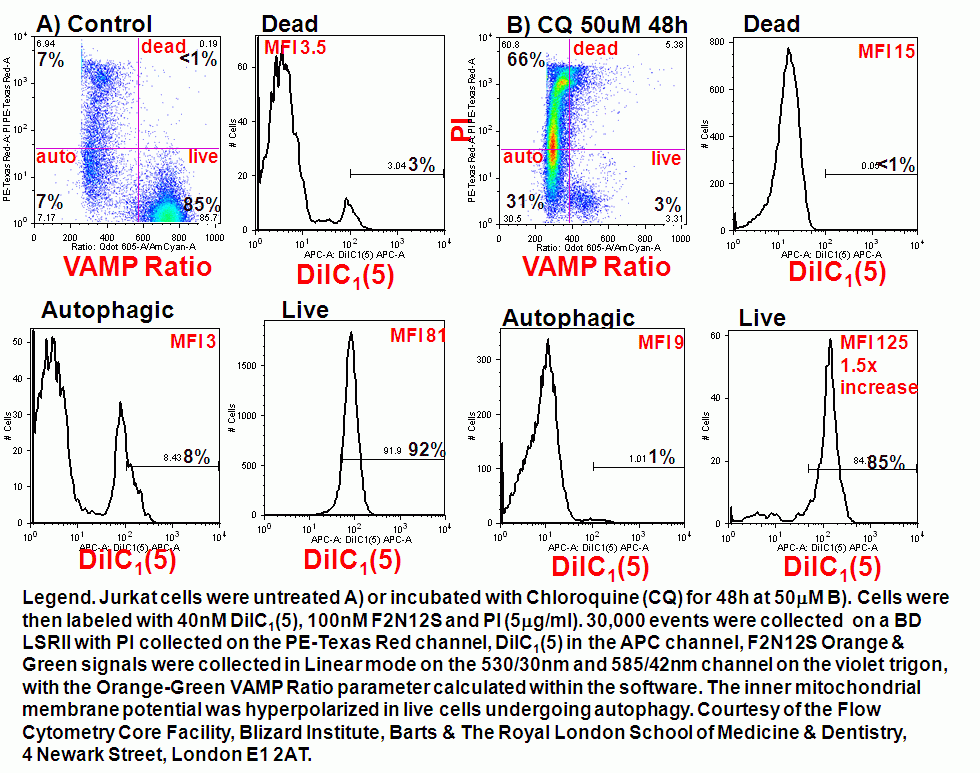
Mitochondria can be degraded when dysfunctional by the process of autophagy or mitophagy, this is a homestatic mechanism in cell maintenaince and survival. However this survival mechanism can result in cell death and can be quantated by measurement of fluorescent carbocyanine dye MitoTracker green which allows the researcher to estimated reductions in cell mitochondrial mass during the process of mitophagy.
During chloroquine induction of autophagy in Jurkat T cells mitochondrial function in live cells show a higher level of inner mitochondrial membrane hyperpolarization indicating a higher energy demand than normal indicating a more rapid production of ATP. Cells labeled with F2N12S membrane asymmetry probe show the presence of live, autophagic and dead cells. The mitochondria of such live cells are show to be hyperpolarized compared to untreated cells, see figure.
Role of the Endoplasmic Reticulum in Autophagic Responses to Drugs
Role of Phospholipids in Autophagic Responses to Drugs
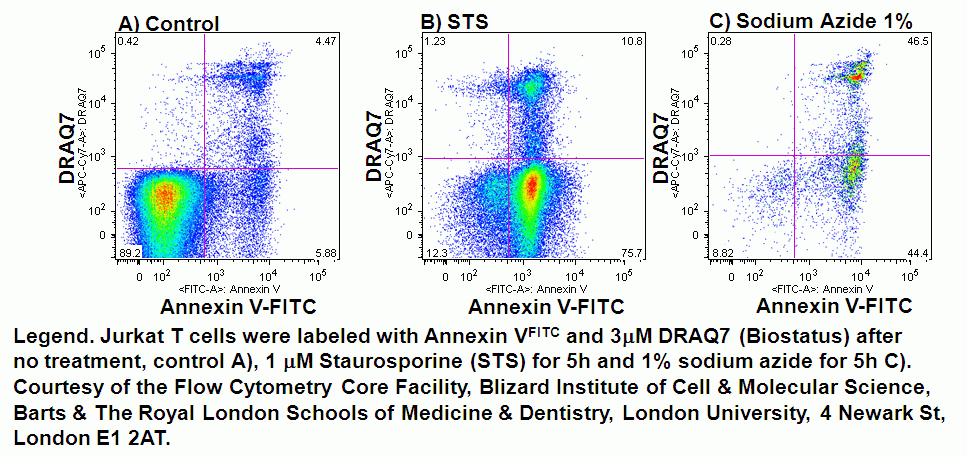
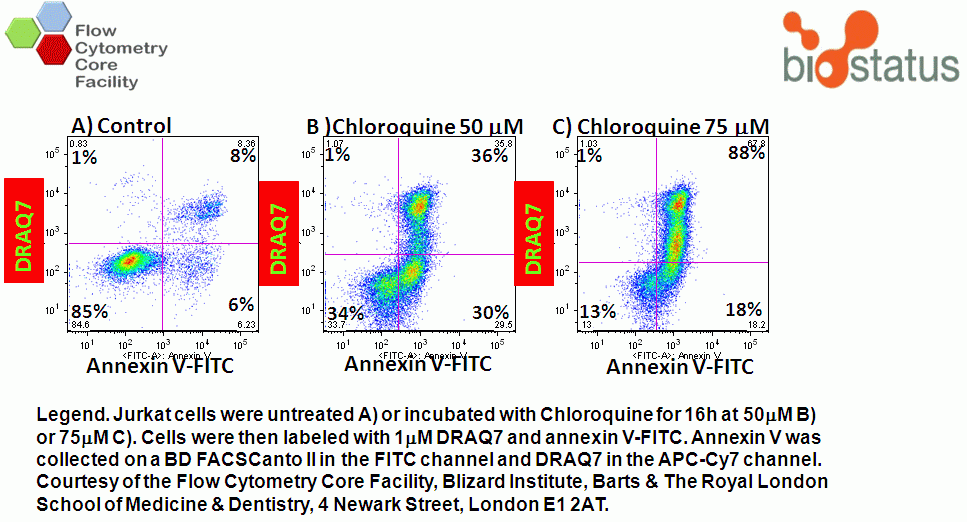
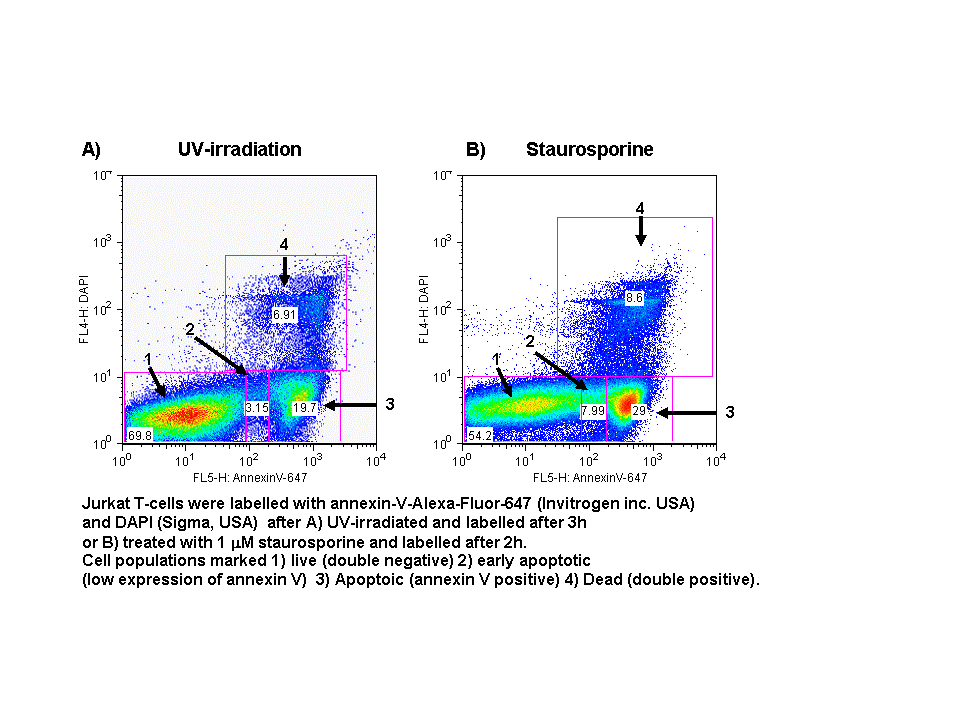
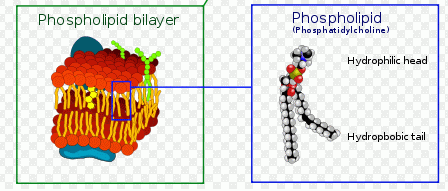
In normal cells, phospholidis, phosphatidylserine (PS) and phosphatidylethanolamine (PE) residues are found in the inner membrane of the cytoplasmic membrane. During autophagy, PE conjugates to microtubule protein LC3B in the autophagosome, see section of LC3B expression. However as in apoptosis the PS residues are translocated in the membrane and are externalised. Annexin-V is a specific PS-PE-binding protein that can be used to detect autophagic cells in an indiscriminate manner by cell surface labeling of such phospholipids. Annexin-V is available conjugated to a number of different fluorochromes including Alexa Fluor-647 and FITC.
Violet ratiometric asymmetry probe
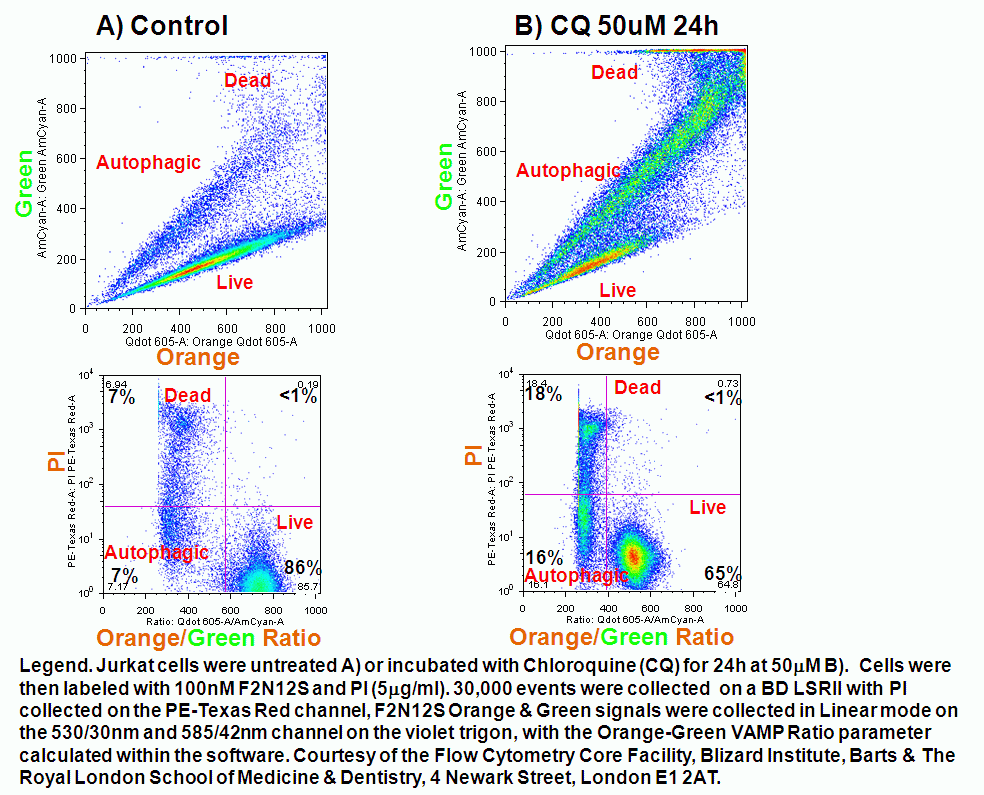
The voltage change associated with the externalization of PS and PE can be detected by plasma membrane probe Bis-oxonol DiBAC4(5) and a new violet ratiometric membrane asymmetry probe, 4'-N,N-diethylamino-6-(N,N,N-dodecyl-methylamino-sulfopropyl)-methyl-3-hydroxyflavone or F2N12S is sensitive to changes in the potential difference or charge on the outer surface of the plasma membrane. The dye has an excited-state of intramolecular proton transfer or ESIPT reaction and in response to changes in the surface charge of the plasma membrane of cells emits fluorescence at two emissions of 530 and 585nm or green and orange fluoresence after excitation with a violet 405nm laser diode. Ratiometric analysis of the 2 emissions with the PMTs switched to linear gives all the advantages of ratiomtric probes over single colour probes in that detected changes are independent of probe concentration, cell size and instrument variation in laser outputs and sensitivity of PMT detectors.
In apoptosis the surface charge of the outer plasma membrane changes which F2N12S can measure by alterations in the intensity of the two emissions, which emit more at 530nm (green) and less at 585nm (orange). Thus ratioing the 585nm emission by the 530nm emission elctronically and plotting this Ratio parameter against cell viability shows the presence of apoptotic cells by a reduced Ratio signal. Dead cells also have a reduced Ratio signal compared to healthy live cells, see figure.
The F2N12S violet ratiometric membrane asymmetry probe is loaded into cells at RT for 5 minutes at 100nM concentration. Cells are not washed after this reaction but must be read within 30 minutes of loading cells. Then annexin V needs to be labeled before F2N12S, if a comparison of the two methods is required. Also although the emission spectra from F2N12S is at 530 and 575nm after 405nm excitation, DAPI with an emission spectra of 461nm cannot be used as a cell viability marker as there appears to be quenching of signals, see figure
Numerous DNA binding viability dyes can be used in the annexin V assay including PI, DAPI and DRAQ7 (Biostatus).
- Model of plasma membrane
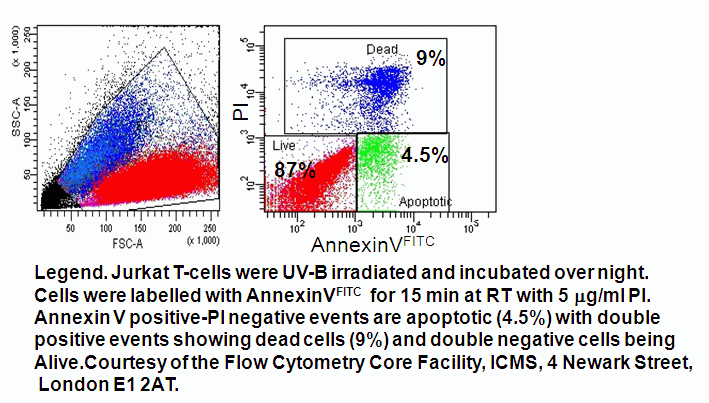
- Quadrant dot plot analysis of peripheral blood mononuclear cells with Annexin V-FITC and PI
- Quadrant dot plot analysis of Jurkat T-cell line with Annexin V-Alexa-Fluor-647 and DAPI
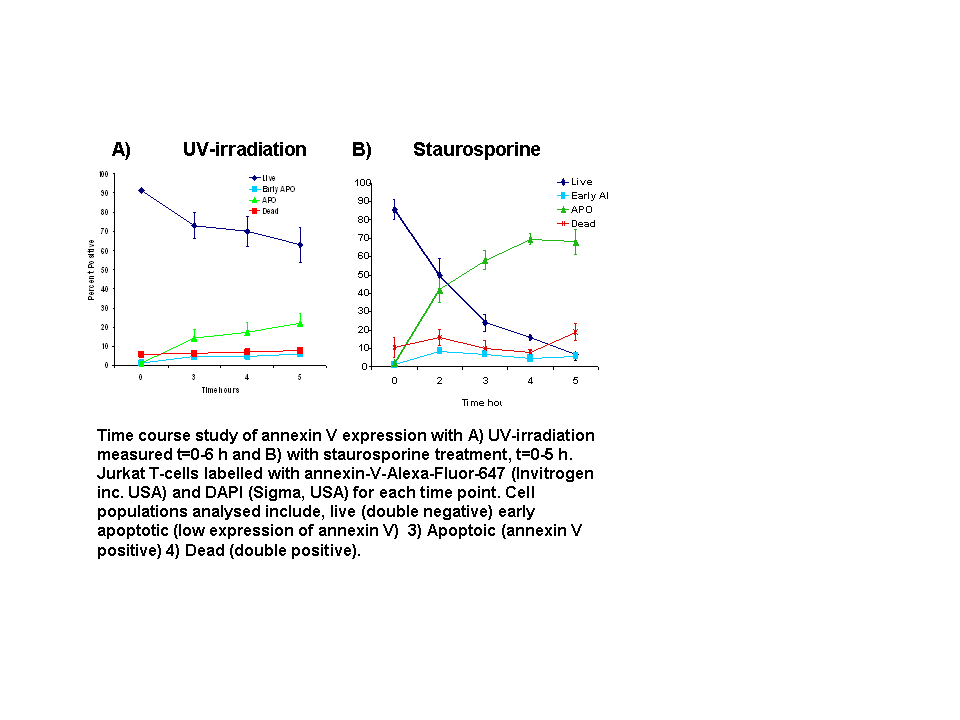
- Annexin V Time Course Study
- Protocol
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}